


Physiological Organ Networks: A Model for Salutogenic Health
Bridging Traditional Vitalism and Modern Systems Science

This is Part 5 in a series discussing herbal medicine in the context of network physiology.
In previous posts in this series, I’ve argued for a shift in how we conceptualize health and disease—moving beyond the mechanical reductionism of what I’ve called Biomedicine 1.0, where the body is treated as a machine made of discrete, replaceable parts. While this model has delivered many lifesaving breakthroughs—particularly in surgery and emergency care—its emphasis on specialization and symptom control has also led to fragmentation, especially in the management of chronic, complex conditions.
In response, I’ve proposed a new framework: Biomedicine 2.0, grounded in systems science and the emerging field of network physiology. This approach reimagines the body not as a collection of isolated organs, but as a dynamic, self-regulating network of signals, rhythms, and feedback loops. It also opens the door to reintegrating the pattern-based insights of traditional medicine within a modern, evidence-informed context.
For many of my readers—especially those trained in naturopathic or functional medicine—this presents both a challenge and an opportunity. At the heart of naturopathic philosophy lies the concept of vis medicatrix naturae—the healing power of nature. Once a cornerstone of a vitalist worldview, the vis today is often invoked more as a poetic or spiritual ideal than a physiological reality, largely disconnected from the language of systems regulation, molecular biology, and clinical science.
Ironically, many “natural” or integrative approaches still operate within the logic of Biomedicine 1.0. Mitochondrial dysfunction, methylation defects, “dirty genes,” environmental toxins, herbal pharmacodynamics—these are real mechanisms, but they are often framed in a reductionist way, treating parts in isolation rather than patterns in context.
If naturopathic medicine is to fully mature—not just as a gentler version of conventional care, but as a paradigm in its own right—it must move beyond what has been called “green allopathy.” This doesn’t require abandoning scientific rigor or reverting to premodern cosmologies. It means evolving our frameworks—just as biomedicine itself is beginning to do—toward models that embrace complexity, coherence, and dynamic regulation.
Here, systems biology and network physiology offer powerful tools for rethinking the vis. Rather than discarding vitalism, we can revitalize it—translating its core insights into the language of emergence, self-organization, and homeodynamic regulation. The organizing forces of life are of a higher order than those governing inert matter—and we now have the conceptual tools to explore them.
In this post, I explore how physiology can be visualized as a network. But rather than focusing on molecular networks of genes, proteins, or metabolites, I propose a clinically relevant model in which organs serve as the nodes, and neuroendocrine-immune signaling between them as the edges. This reframing allows us to assess functional coherence across the system—not just biochemical status within it.
To do this well, however, we must distinguish between organs as anatomical structures and organs as dynamic, functional processes.
From this perspective, any disease state can be understood as a disruption in network coherence—where one or more organ “nodes” fail to regulate effectively. Treatment, then, becomes less about suppressing symptoms or boosting isolated pathways, and more about restoring the relational dynamics that sustain system-wide balance.
Such thinking invites us to move beyond supplement stacks and toward interventions—like polyherbal formulations—that restore pattern and coherence across the entire network.
In short, this is a call to rethink the vis—not as a relic of metaphysical vitalism, but as an emergent property of a complex, self-organizing living system—one we can now begin to observe, map, and support with increasing clarity.

Reframing Physiology: Organs as the Hubs of Healing
Network science has revolutionized how we understand complexity in biology. Instead of analyzing isolated parts, systems biology constructs biological networks—maps that reveal how genes, proteins, and metabolic pathways interact in dynamic, context-sensitive ways. This shift from linear causality to interconnected regulation has reshaped fields ranging from oncology to pharmacology.
Take gene regulation. Rather than targeting a single gene, systems biologists examine expression data, protein-binding assays, and perturbation experiments to uncover functional relationships. These are visualized as graphs: nodes (e.g., genes or proteins) connected by edges (e.g., regulatory or physical interactions). Within these protein-protein interaction (PPI) networks, certain nodes emerge as hubs—highly connected regulators of metabolism, stress response, or cell fate.1 Recognizing these hubs has transformed our understanding of disease mechanisms and helped identify strategic points of therapeutic intervention.
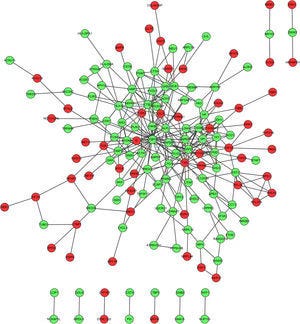
Visualizing Gene Activity in Asthmatic Children
This image shows a protein–protein interaction (PPI) network constructed from microarray gene expression data in children with asthma. Each dot represents a protein encoded by a gene, with red nodes indicating upregulated genes(increased activity) and green nodes indicating downregulated genes (decreased activity). The lines between them represent known or predicted interactions, helping to visualize how changes in gene activity may influence broader biological pathways and systems.2
This same network logic can be extended to understanding polyherbal medicine, especially traditional formulas from systems like Traditional Chinese Medicine (TCM). Advances in phytochemistry and bioinformatics now allow us to map herbs to their bioactive compounds and known molecular targets. These targets can be layered onto existing interaction networks, generating herb-compound-target-disease (HCTD) models. Such models help visualize how complex formulas modulate physiology across multiple nodes and pathways. (I've created one such example of a network pharmacology-based analysis for Chinese herbs used in migraine, viewable here.)
Yet as elegant as these molecular maps are, they remain largely academic. Most clinicians lack access to real-time omics data, and more importantly, traditional medicine has never operated at the molecular level. It has always operated through the lens of organs.
Organs serve as the phenomenological scaffolding for both diagnosis and treatment. In TCM, a pattern like Liver heat emerges from a recognizable constellation of symptoms, pulse and tongue findings, and clinical experience. In biomedicine, liver dysfunction is evaluated through laboratory markers, imaging, and histopathology. While the language differs, both systems treat the organ as a functional unit—a focal point where systemic imbalance becomes visible, interpretable, and actionable.
This insight offers a clinically grounded bridge to network medicine: one in which organs are the nodes, and neuroendocrine, autonomic, and immune signaling between them forms the edges. Rather than building from molecules upward, we can construct networks from the top down—from whole systems grounded in embodied experience. Physiological relationships like the gut-brain axis, kidney-heart interaction, or liver-adipose crosstalk reflect real, bidirectional coupling—mediated through shared signaling pathways, rhythmic entrainment, and metabolic interdependence.
Emerging computational methods, such as parenclitic network analysis, allow us to model these inter-organ relationships using clinical data. Parameters like lab values, HRV metrics, cytokine profiles, and sleep patterns can be used to construct individualized organ networks. Graph theory tools—centrality, path length, modularity—can then identify weak links, overloaded hubs, or areas of dysregulation within a person’s physiological network.
This opens powerful new frontiers for clinical insight. By mapping how regulation flows through an individual's organ network—via stress signaling, autonomic tone, or inflammatory feedback—we gain a roadmap for targeted intervention. Treatments, including herbal formulations, can be selected not just for symptom relief, but to restore network coherence: reinforcing weak nodes, rerouting flows, and stabilizing disordered patterns.
This is the promise of a network-informed physiology: not just a new model of the body, but a new way of thinking, seeing, and healing—one that is at once modern and deeply aligned with traditional principles.
The Threefold Organ: From Anatomy to Dynamic Function
If organs are to serve as the nodes in our physiological network, we must first ask: What is an organ? In both traditional medical systems and modern systems biology, organs are not merely anatomical structures but dynamic processes—coordinating diverse physiological functions across time and space. An organ is not simply a localized piece of tissue, but a functional entity: a node of integration that emerges from complex, often spatially distributed activity.
To reflect this layered reality, I use capital letters (e.g., Liver, Kidney, Gallbladder) to refer to the functional Organ—as conceptualized in traditional and systems-based models—and lowercase (e.g., liver, kidney) to refer strictly to anatomical structures.
This distinction is neither symbolic nor arbitrary—it reflects the fundamental nature of living systems. Unlike machines, which are constructed from discrete parts, organisms develop from an integrated whole. Life does not arise through mechanical assembly, but through processes of division, differentiation, and patterned emergence. From a single fertilized egg, complexity unfolds organically, following a developmental logic that is radically different from stacking Legos, parts, or even cells together.
In the unfolding process of embryogenesis, polarity soon emerges—typically expressed as a dynamic tension between neuro-sensory and metabolic activity. These opposing forces are mediated by a third, harmonizing function: rhythm. This threefold logic of life becomes discernible as early as the third week of development.
The human form reflects this triadic architecture. The three embryonic germ layers—ectoderm, mesoderm, and endoderm—give rise to distinct yet interdependent physiological systems:
Ectoderm → Neuro-sensory system
Mesoderm → Rhythmic (circulatory-muscular) system
Endoderm → Metabolic-digestive system
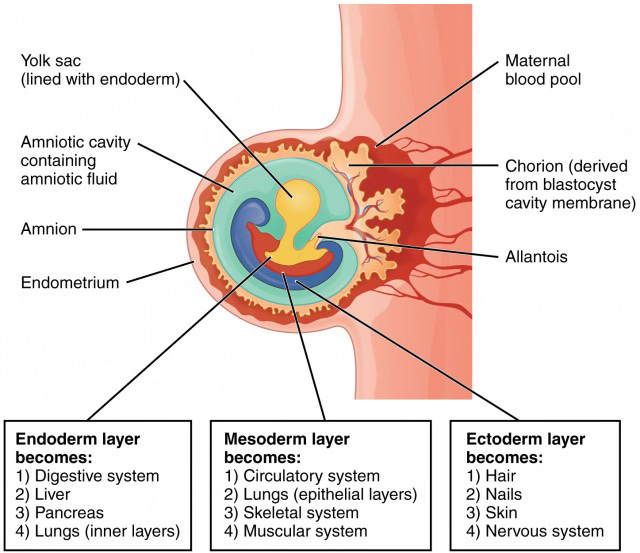
Gastrulation: Week 3 of human embryological development3
These are more than anatomical origins; they define functional domains. And this triadic logic recurs at multiple scales of biology. For example, within the cell:
The cell membrane acts as the nerve-sensory interface—detecting, receiving, and relaying signals.
The nucleus, endoplasmic reticulum, and ribosomes serve as metabolic centers—producing and regulating proteins and genetic material.
The cytoplasm and mitochondria carry out rhythmic and respiratory functions—cycling energy and maintaining intracellular coherence.
This brings us to a critical point: the laws governing living systems differ qualitatively from those of inert matter. We can kill an organism, dissect it, and analyze its parts—but in doing so, we destroy the very dynamics we seek to understand. Life is not merely the sum of its components, but the expression of relationships: temporal, functional, and emergent.
Indeed, when we observe how living systems grow, repair, and self-regulate, we find ourselves naturally reaching for more fluid, relational terms: anabolism and catabolism, incretion and excretion, sclerosis and inflammation. We don’t merely think in mechanisms—we think in polarities and processes. This is not imprecision; it’s fidelity to life’s complexity.
Thus, the threefold logic of function—nerve-sensory, metabolic-digestive, and rhythmic-circulatory-breathing processes—is not a projection, but an emergent feature of life itself. When applied to organs, this framework invites a shift: from a static map of anatomical parts to a fluid physiology of patterned interaction. Organs become less things, and more processes embedded within networks—shaped by, and participating in, broader flows of regulation.
This reframing creates a vital bridge between traditional organ-pattern reasoning and modern systems biology. It gives us a common language to talk about terrain, coherence, and imbalance—in ways that are both scientifically grounded and clinically intuitive.
And it’s this language that will be essential for the network medicine of the future.
A Philosophical Digression: From Matter to Form to Function
To deepen our understanding of the organ as a dynamic process—not merely a structure—it helps to briefly step into the history of ideas.
In classical philosophy, Aristotle offered a foundational framework for understanding living beings through what he called hylomorphism: the view that all natural things are composed of both matter (hyle) and form (morphe).4 Matter provides the substrate—the potentiality—while form is the active principle that gives shape, structure, and function. Importantly, form is not just an abstract idea; it is a real, formative impulse that organizes substance into coherent living beings.
Later thinkers, including the Austrian philosopher Rudolf Steiner (1861-1925), extended this concept.5 For Steiner, as for some modern systems theorists, form is not imposed from outside nor reducible to material causation. Instead, it represents a kind of informational blueprint—an inner principle of organization that guides growth, differentiation, and coherence in living systems.
Aristotle used the term entelechy to describe this inner formative drive—the realization of a being’s inherent potential. In the early 20th century, biologist Hans Driesch (1867-1941), a neo-vitalist, proposed that embryological development was governed not by mechanical forces alone, but by an organizing field—a non-material principle guiding morphogenesis.6 Though controversial in scientific circles, this idea anticipated modern theories of morphogenetic fields and biofields, which suggest that information—rather than mere chemistry—plays a central role in biological development and regulation.
This concept resonates deeply with many complementary and traditional medical systems. When practitioners speak of “energy” or “energetic organs,” they are often referring—consciously or not—not to measurable physical energy (like electromagnetism or ATP), but to something more like entelechy: an intrinsic organizing intelligence within the system. This informational quality is what shapes the flow of energy and matter, not the other way around.
To clarify this further, consider a simple analogy: a traffic light. The electrical energy required to change a red light to green is minimal. Yet that small signal informs the flow of tremendous kinetic energy—cars and trucks barreling down a road. It’s not the amount of energy that matters, but how that energy is directed by information.
We can extend this analogy to the human body. Think of matter and energy—genes, proteins, hormones, cells, and anatomical organs—as the hardware of the system. The patterns of coordination between them—the signaling, feedback, and regulation—function as the software. The “organ” in the anatomical sense is hardware. The organ as process, the functional Organ, is software: a set of informational relationships that guide how the parts behave in context.
This distinction matters. Too often, complementary medicine still speaks of “energetic organs” in a way that feels out of step with contemporary science. But if we instead speak of informational organs—the organ’s entelechy, its formative logic—we begin to bridge traditional insight with systems biology. We are no longer invoking vague energy fields, but rather patterned informational flows: the software of living systems that regulates, coordinates, and adapts.
Here, network science offers a crucial conceptual advance. In a biological network, the nodes represent things—genes, cells, organs. The edges represent relationships—signaling pathways, feedback loops, regulatory rhythms. If we temporarily imagine removing the nodes and focus only on the edges, what remains is not emptiness, but a field of interaction. This field is not a substance but a pattern—a living matrix of dynamic relationships through which regulation and coherence emerge.
From this perspective, what traditional medicine calls “energy” and what Aristotle called entelechy may now be interpreted as emergent relational fields—the informational architecture that shapes and coordinates life. These fields are not metaphysical speculations; they are observable in the form of autonomic tone, cytokine crosstalk, hormonal entrainment, and bioelectrical gradients. They are the edges of the physiological network in action.
In this view, an organ is not just a thing, but a pattern—a process that arises within and contributes to a larger field of communication. This is what we mean by the organ as process, and why network physiology may finally give us the tools to study what traditional systems have long intuited: that life flows not only through parts, but through the relationships between them.
The Liver–Gallbladder (LG) Node: Threefold Function and Central Regulation
Let’s consider the Liver as a case study—arguably the most multifaceted organ in the human body. Within a systems framework, the Liver is not merely a metabolic hub, but a regulatory node, maintaining coherence between the organism’s internal milieu and its external environment. It mediates critical domains: stress response, detoxification, digestion, immune surveillance, and reproductive timing. This networked intelligence becomes apparent when we examine how the Liver integrates neural, hormonal, immune, and metabolic signals.
Anatomically, the liver consists of hepatocytes and specialized cells arranged within a vascular lattice optimized for transformation, exchange, and clearance. But functionally, the Liver extends well beyond its gross morphology. It acts as a distributed system, modulating systemic levels of glucose, IGF-1, bile acids, amino acids, and acute-phase proteins. Its influence radiates across multiple tissues via feedback loops and signaling gradients.
If we view the Liver as a node, we discover within it a micro-network that expresses a threefold functional polarity:
Neuro-sensory domain: This includes both afferent and efferent autonomic fibers (sympathetic and parasympathetic), which regulate hepatic tone and responsiveness. But it also extends to endocrine signaling (e.g., hepatokines like IGF-1 and FGF21) and immune communication (e.g., cytokines from Kupffer cells). These sensory and signaling elements together form a perceptual and coordinating layer within the node—orienting the liver to internal and external cues.
Metabolic domain: The Liver performs both incretory (anabolic) and excretory (catabolic) functions.
Incretory: It builds and nourishes—synthesizing glucose (via gluconeogenesis), plasma proteins, glutathione, and lipoproteins. These processes occur primarily within hepatocytes, as blood flows inward from the portal triads toward the central veins.
Excretory: Simultaneously, the Liver clears and detoxifies—conjugating hormones, neutralizing xenobiotics, and producing bile. While the gallbladder stores and concentrates bile, the Gallbladder as a functional entity expresses the Liver’s excretory rhythm. Even after cholecystectomy, this rhythmic bile secretion persists—reminding us that function precedes form.
Rhythmic-regulatory domain: These opposing poles—building and clearing—are entrained to circadian and ultradian rhythms. Nutrient sensors, hepatic clock genes, and autonomic tone coordinate the timing of glucose release, bile production, and hormonal responsiveness. The Liver thus expresses a temporal intelligence—regulating not only what happens, but when.
The Liver and Gallbladder together form the Liver–Gallbladder (LG) Node—a functional unit defined less by anatomical proximity than by their dynamic polarity. The Liver, solid and nourishing (yin), orchestrates transformation, growth, and anabolic processes—what we might call incretory activity. The Gallbladder, hollow and decisive (yang), facilitates release, direction, and excretory function. In this framing, “Liver” represents the nerve-sensory, metabolic, and rhythmic forces that support inward-building and patterning; “Gallbladder,” those that enable outward-flowing, clearing, and resolution.
These polar functions are not separate, but braided through a rhythmic exchange—most visibly in the sinusoidal circulation of blood, bile, and lymph. This flow bridges center and periphery, nourishing and detoxifying, organizing and unblocking. Through this oscillating process, the hepatobiliary system expresses itself not as two organs, but as a single, intelligent node within the broader physiological network—a hub of coherence, communication, and transformation.
The LG Node and Cross-System Integration
The LG Node participates in several cross-system axes, forming a lattice of inter-nodal communication. Some examples include:
Liver–Hypothalamus–Pituitary Axis: The Liver plays a pivotal role in the growth axis. Pituitary-derived growth hormone (GH) stimulates hepatic production of IGF-1, which promotes anabolic processes in muscle, bone, and connective tissue. This constitutes a bidirectional feedback loop between neuroendocrine command and hepatic execution.
Liver–Muscle Connection: During exertion, skeletal muscle releases lactate, which the Liver converts back to glucose via the Cori cycle. This loop is not only metabolic but rhythmic, aligning peripheral demand with central supply in a dynamic feedback circuit.
Liver–Adipose–Brain Axis: Hepatokines such as FGF21 influence appetite, thermogenesis, and insulin sensitivity—linking hepatic state to hypothalamic signaling and adipose metabolism. Conversely, chronic hepatic inflammation can disrupt autonomic tone, contributing to fatigue, mood dysregulation, and circadian disturbance.
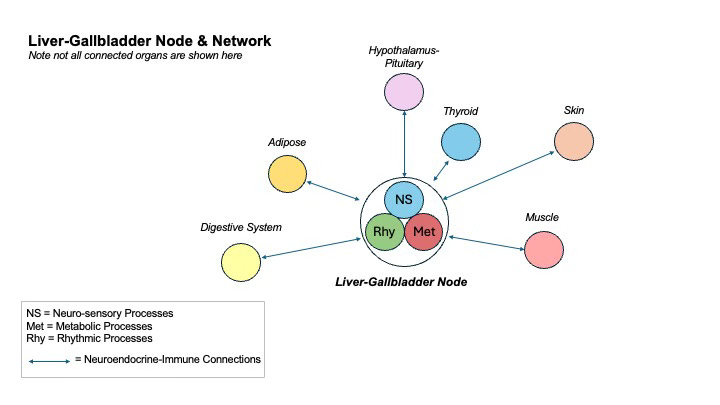
The LG Node as Regulator of Life Processes
Far from serving merely as a passive filter or biochemical processor, the Liver–Gallbladder (LG) Node functions as a dynamic regulator of systemic life processes. It governs the quality and composition of blood, orchestrates the detoxification and clearance of endogenous and exogenous compounds, and plays a critical signaling role—modulating hormonal rhythms, metabolic tone, and energetic availability in tissues such as muscle and adipose. Through these interwoven activities, the LG Node contributes to the organism’s core regulatory integrity: sustaining anabolism where needed, catalyzing catabolism when appropriate, and maintaining homeodynamic balance between storage and mobilization.
Conclusion: From Organs to Patterns—Toward a Truly Salutogenic Practice
As we’ve seen, a network-based view of physiology requires us to rethink the nature of organs themselves—not as fixed anatomical structures, but as dynamic processes shaped by flows of information. The true “organ” is not just tissue, gland, or cell, but the entelechy that coordinates their function: a pattern-forming field, an emergent informational coherence that guides physiological activity. What traditional systems once called “energetic” organs may be better understood, in today’s scientific language, as informational nodes embedded in a dynamic network.
Network science now offers the tools to map this informational terrain. If we abstract away the nodes—cells, genes, hormones—and focus on the edges, the flows between them, we begin to perceive a field: the emergent dynamics of signaling, regulation, and interaction. This is where the organ-process lives—not in the matter itself, but in its self-organizing, relational dance. It is this field of interaction that becomes disordered in illness and coherent in health.
In future posts, I will explore each of the major functional organ nodes—Renal-Urogenital, Gastrointestinal, Cardiovascular, and Respiratory—and show how each operates not only anatomically, but as a coherent signaling unit with its own tendencies toward imbalance and self-correction.
Building on this, we will look at the idea of organ terrains—stable patterns or "attractor states" that arise when organ-network dynamics become distorted. These terrains, long recognized in traditional systems (as "stagnation," "heat," "deficiency," etc.), are not merely symptoms, but signatures of disrupted entelechy—disordered patterns in the informational field.
A truly salutogenic approach to medicine must begin here: by identifying the weakest organ-process in the system and determining the terrain in which it is trapped. Only then can we intervene meaningfully—not with blunt-force pharmaceuticals or isolated nutraceuticals, but with patterned information capable of reshaping the distorted field.
This is where polyherbal intelligence offers its great promise. Traditional formulas weren’t just collections of active compounds; they were coherent signals, crafted over centuries, capable of nudging an organ-network back into coherence. Their intelligence lies not in the potency of any single constituent, but in their capacity to communicate with the body in its own language: that of pattern, process, and form.
In a world of fragmented systems and reductionist solutions, we are called to remember the wisdom of pattern and the power of form. The organ is not just a thing, but a process. And healing, in its truest sense, is the restoration of meaningful form.
Note: Some images in this post were generated with the assistance of an AI model.
Safari-Alighiarloo N, Taghizadeh M, Rezaei-Tavirani M, Goliaei B, Peyvandi AA. Protein-protein interaction networks (PPI) and complex diseases. Gastroenterol Hepatol Bed Bench. 2014 Winter;7(1):17-31. PMID: 25436094; PMCID: PMC4017556.
Chen Y, Qiao J. Protein-protein interaction network analysis and identifying regulation microRNAs in asthmatic children. Allergol Immunopathol (Madr). 2015 Nov-Dec;43(6):584-92. doi: 10.1016/j.aller.2015.01.008. Epub 2015 May 12. PMID: 25979194.
Embryonic Development | Anatomy and Physiology II. (n.d.). https://courses.lumenlearning.com/suny-ap2/chapter/embryonic-development/
The Editors of Encyclopaedia Britannica. (1998, July 20). Hylomorphism | Form, matter & Prime Mover. Encyclopedia Britannica. https://www.britannica.com/topic/hylomorphism
Contents - GA 9. Theosophy (1971) - Rudolf Steiner Archive. (n.d.). https://rsarchive.org/Books/GA009/English/AP1971/GA009_index.html
Driesch, H. (2020). The science and philosophy of the organism. BoD – Books on Demand.