
The Pathway Is a Waveform
The hidden geometry of biochemical pathways and the limits of nutrient-based medicine


Introduction
Open any biochemistry textbook and you will find the same image: a network of boxes and arrows. Molecules are nodes; enzymatic reactions are edges. The citric acid cycle appears as a tidy wheel, glycolysis as a ladder, the electron transport chain as a relay. These diagrams have been extraordinarily productive. They gave us the logic of metabolic disease, the rationale for nutritional supplementation, the targets of most pharmaceutical drugs. They remain essential teaching tools.
These network-like images are also, in a precise sense, projections: shadows cast by a higher-dimensional reality onto a two-dimensional page. Taken at face value, they suggest that the human organism is essentially a bag of chemicals, and that health is a matter of adding the missing ones or removing the excess. In the framework developed in this series, what casts these shadows are the counterspatial morphogenetic fields whose phase-field geometry organizes the molecular pattern from a deeper level of reality. The shadow is real, but the field is primary.
This post examines what the shadow leaves out. Over the past three decades, converging lines of evidence from enzymology, structural biology, quantum biology, and biophysics have revealed that cellular biochemistry has an underlying geometry—spatial, dynamic, and coherent—that the nodes-and-edges picture systematically obscures.
Metabolic enzymes do not float independently in a well-stirred solution, waiting for substrates to diffuse into their active sites. They assemble into precisely organized complexes, channel intermediates directly without releasing them to bulk solution, and catalyze reactions through quantum-mechanical processes that depend on the coordinated dynamics of the whole assembly.1 The medium in which all of this occurs, cellular water, is not a passive solvent but a structured, oscillatory field that threads through and between these complexes.2
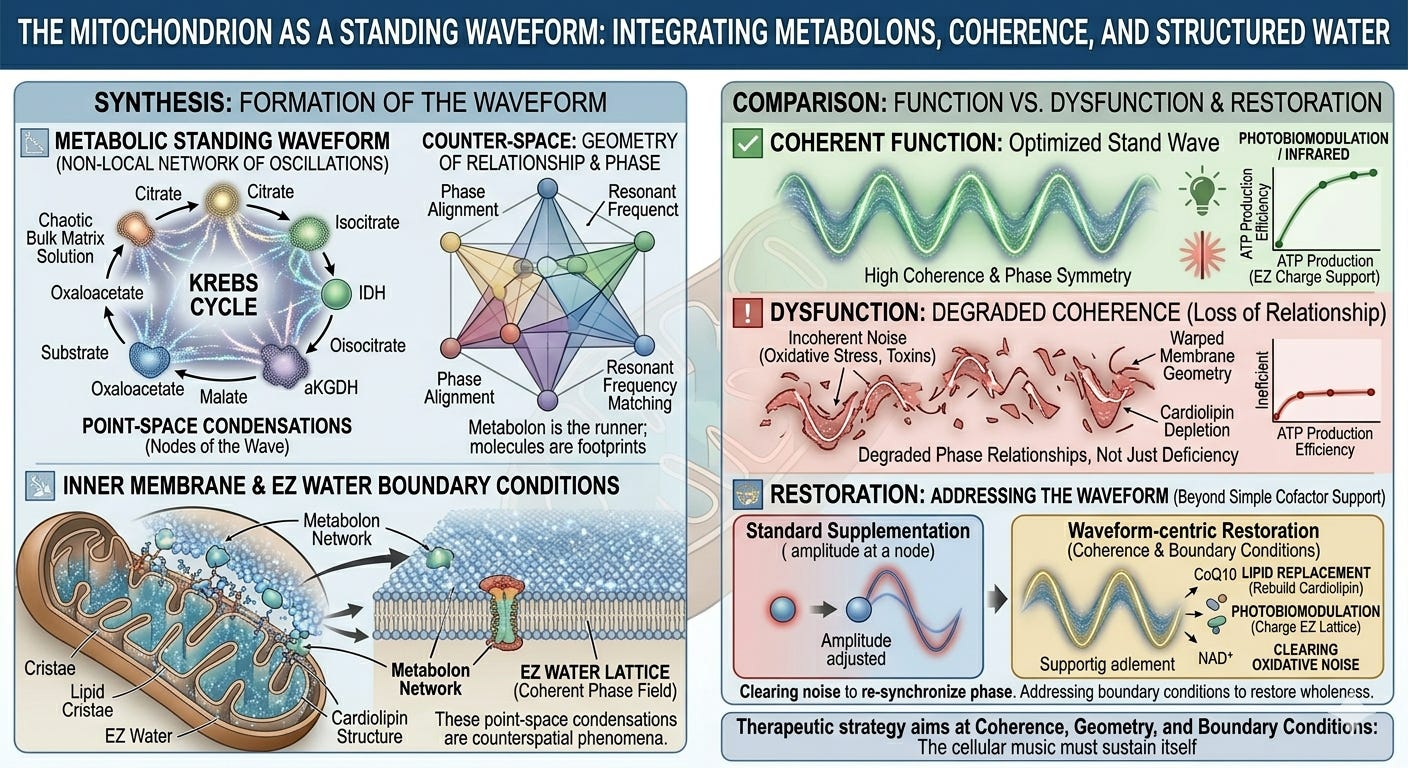
When this evidence is taken seriously, a new picture emerges: not a pipe network to be pressure-fed, but a standing waveform to be tuned. The therapeutic approaches common in functional medicine and naturopathy—high-dose cofactors, targeted substrate loading, pathway-specific nutrient protocols—are not wrong so much as incomplete: they address the point-space condensation while remaining blind to the waveform that organizes it. They supply notes without attending to the resonance of the instrument.
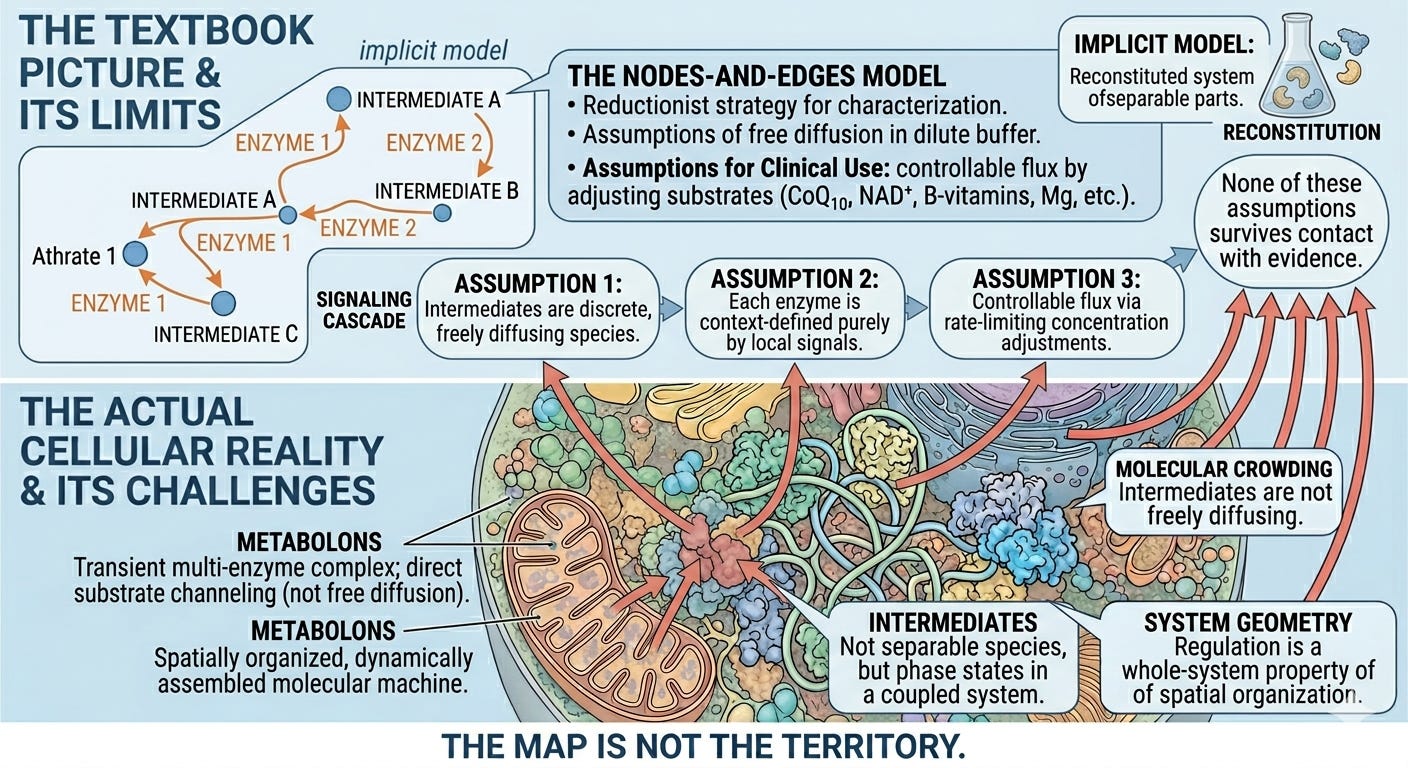
The Limits of Biochemical Network Models
The nodes-and-edges model of metabolism has a specific intellectual history. It emerged from the reductionist program of twentieth-century biochemistry, which achieved its greatest triumphs by isolating enzymes, characterizing their kinetics in dilute solution, and then reconstituting pathways from purified components. This was the right method for the questions being asked. Krebs could not have discovered the citric acid cycle by studying intact mitochondria; the cycle had to be broken apart first.3
The problem is that what works as a discovery strategy becomes, over time, an ontological assumption. Having learned the cycle by taking it apart, biochemistry came to think of it as a collection of separable parts. The reconstituted system—enzymes in dilute buffer, intermediates freely diffusing—became the implicit model of what was happening in the cell. The map displaced the territory.
Three assumptions follow from this displacement, each clinically consequential.
Intermediates are assumed to exist as discrete, freely diffusing molecular species between reactions.
Each enzyme is assumed to operate in a context defined entirely by local chemical signals—substrate concentration, cofactor availability, allosteric state, and, in more sophisticated models, the hormonal and inflammatory environment conveyed by signaling pathways. This is a genuine advance over simple Michaelis-Menten kinetics; it acknowledges that the cell is not a test tube. But signaling pathways are themselves represented as nodes and edges—receptors, second messengers, kinase cascades, transcription factors connected by arrows. The wiring diagram grows more complex; its ontology does not change. Regulation is still conceived as one molecular species acting on another, never as a property of the geometric and coherent organization of the system as a whole.
Pathway flux is therefore controllable by adjusting the concentration of rate-limiting substrates, cofactors, or signaling molecules, which is the logic behind the widespread clinical use of CoQ10, NAD+ precursors, B-vitamin complexes, magnesium, and anti-inflammatory agents.4
None of these assumptions survives close contact with the actual cellular evidence.
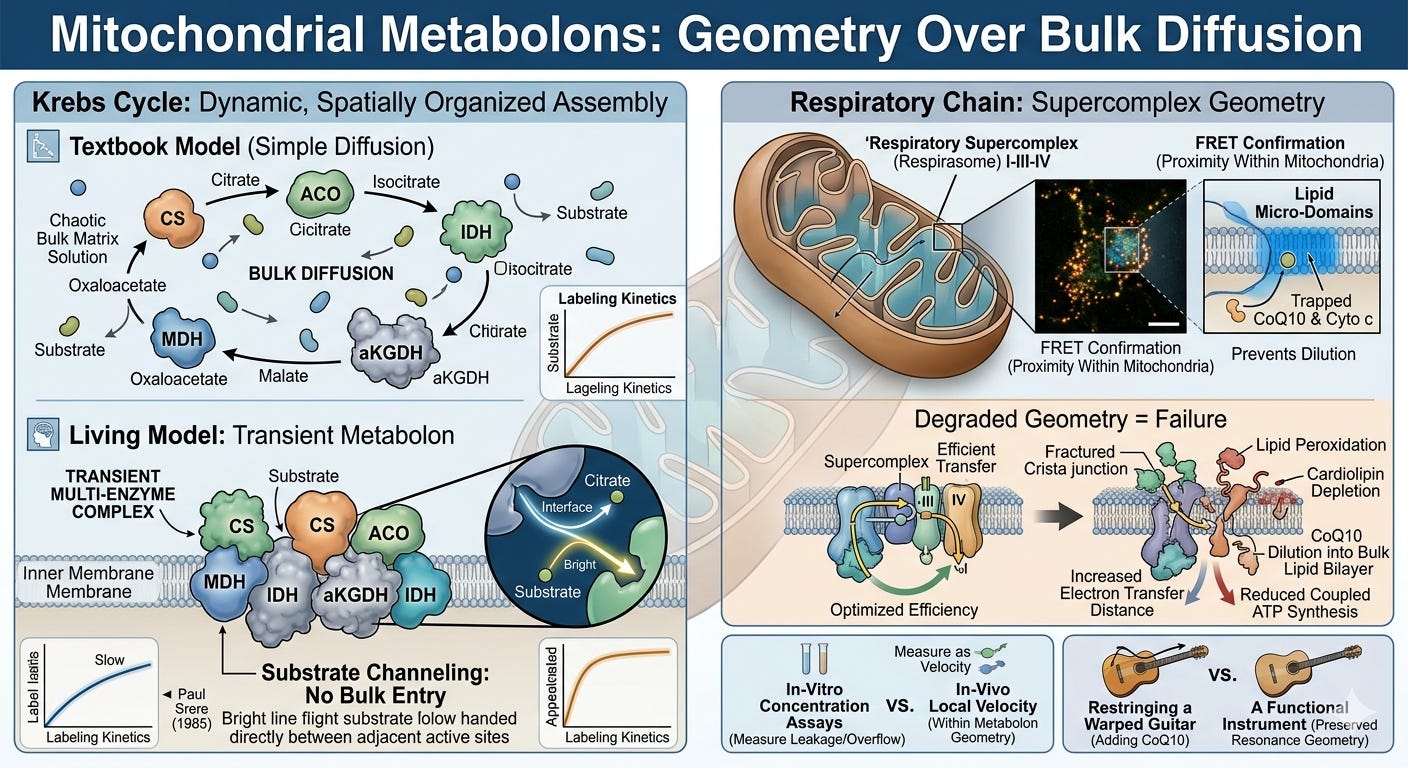
Metabolons: The Geometry of Biochemistry
The first major departure from the textbook picture is the metabolon: a transiently assembled multi-enzyme complex in which substrates are channeled directly from one active site to the next without diffusing into bulk solution. The concept was formalized by Paul Srere in 19855, and evidence for metabolons in the citric acid cycle has accumulated steadily since.
Co-immunoprecipitation experiments pull down multiple Krebs cycle enzymes together under near-physiological conditions—citrate synthase, aconitase, isocitrate dehydrogenase, α-ketoglutarate dehydrogenase, and malate dehydrogenase associate in complexes disrupted only under denaturing conditions.6 FRET studies confirm that these enzymes maintain close proximity within intact mitochondria.7 Kinetic isotope tracing demonstrates that labeled intermediates appear at the next enzymatic step faster than free diffusion through bulk solution could account for—the intermediate never enters bulk solution at all.8
What this means structurally is that the Krebs cycle in the living mitochondrion is not a wheel of reactions connected by diffusion. It is a spatially organized, dynamically assembled molecular machine in which the geometry of assembly is as functionally important as the chemistry of each individual step. The intermediates—citrate, isocitrate, α-ketoglutarate, succinate, fumarate, malate, oxaloacetate—are less like independent molecular commodities and more like transient phase states in a highly coherent, coupled system. While minor fractions of these metabolites can escape into bulk solution to act as secondary signaling messengers or biosynthetic precursors, the primary catalytic flux driving ATP production occurs within this closed circuit.
Consequently, their concentrations in bulk solution, which is what standard organic acid assays measure, represent the leakage or the overflow, not the actual localized velocity inside the metabolon.
This is a decisive challenge to simple supplementation logic. Because oxaloacetate is directly tunneled from malate dehydrogenase to citrate synthase, flooding the bulk matrix solution with exogenous oxaloacetate cannot directly accelerate that closed-loop transition. It forces a complex, system-wide equilibrium shift at the margins, but it completely bypasses the real architectural bottleneck: the assembly status of the machine itself.
The respiratory chain extends this picture further. Complexes I, III, and IV of the electron transport chain assemble into supercomplexes known as respirasomes, with precise stoichiometry and geometry conserved across species.9 This structural configuration creates dedicated lipid micro-domains that trap electron carriers like CoQ10 and Cytochrome c, preventing them from diluting into the wider membrane.
Thus, the geometry of the supercomplex is not incidental to its function; it is the absolute precondition for it. Disrupt this geometry through, say, membrane lipid peroxidation, cristae remodeling, or cardiolipin depletion, and these micro-domains dissolve. CoQ10 dilutes into the bulk lipid bilayer, electron transfer distance increases exponentially, efficiency drops, reactive oxygen species production rises, and coupled ATP synthesis is impaired.10
The system fails not because a part is missing, but because the spatial organization that makes its function possible has been degraded.
This concept has important implications for mitochondrial support protocols. The standard rationale for nutrients such as riboflavin, thiamine, lipoic acid, and CoQ10 is that they serve as cofactors or electron carriers at specific enzymatic steps, replacing missing parts and restoring the flux. This logic is not false; genuine cofactor deficiencies are real and correctable. But it addresses the chemistry of individual nodes while leaving the geometry of the system untouched.
If the supercomplex has unraveled due to cardiolipin peroxidation, if the cristae junctions have collapsed under chronic inflammatory signaling, or if the interfacial EZ water layer protecting the matrix has lost its charge separation, then an exogenous electron carrier has no coherent topology to navigate. Supplying CoQ10 to a geometrically degraded respiratory chain is like restringing a guitar whose body has warped: the string can be tightened, but the instrument has lost its structural capacity to hold a standing wave and sustain resonance.
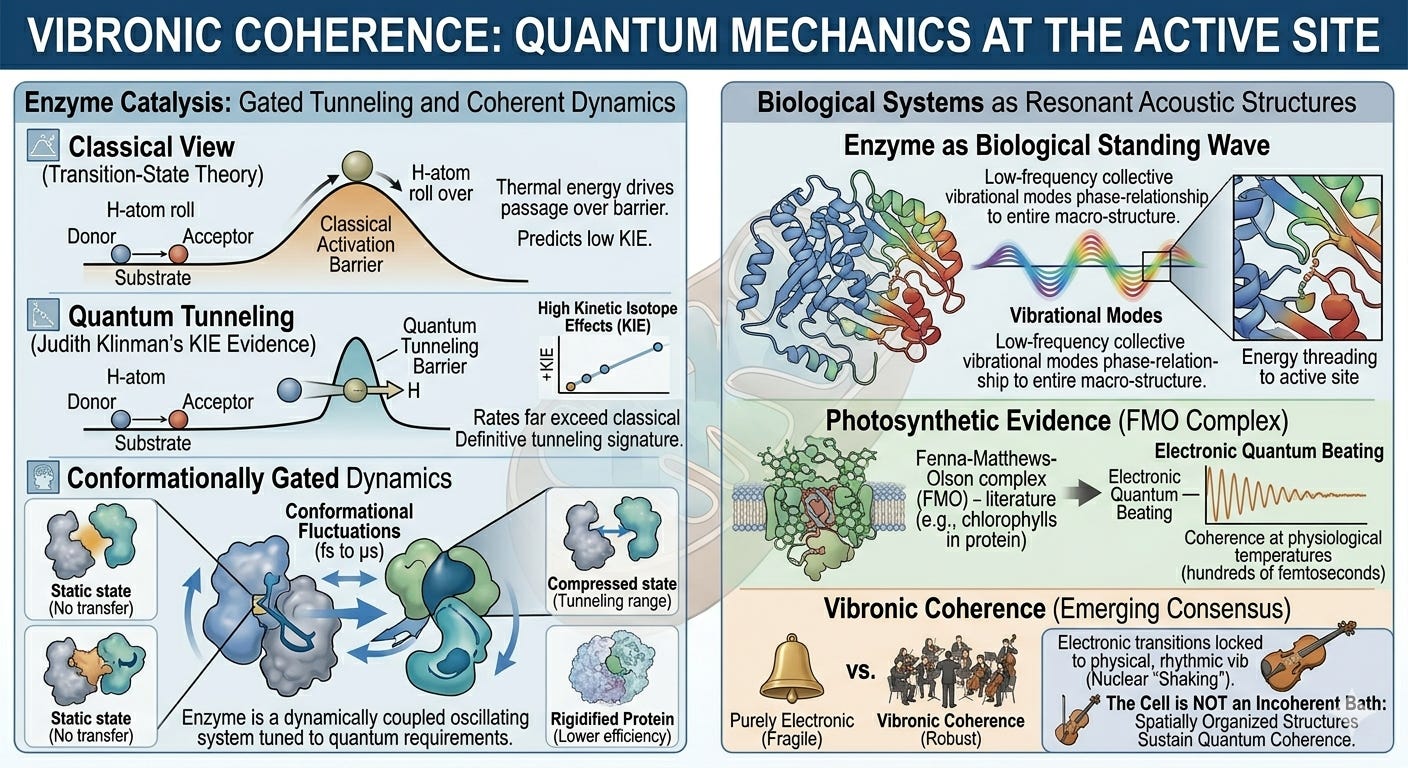
Vibronic Coherence: Quantum Mechanics at the Active Site
The metabolon evidence establishes that pathway geometry matters. The next layer of evidence establishes that the dynamics of that geometry, its oscillatory, wave-like character, are not incidental noise but central to catalytic function.
The most extensively studied case is hydrogen and proton tunneling in oxidoreductase enzymes. Classical transition-state theory predicts that chemical bonds break and form when ambient thermal energy drives reactants over a fixed activation energy barrier. For a wide class of enzymes, this prediction fails quantitatively. Judith Klinman’s group, working on alcohol dehydrogenase and other enzymes, demonstrated that hydrogen transfer rates show kinetic isotope effects far exceeding classical predictions: a definitive signature of quantum tunneling through the energy barrier rather than physical passage over it.11 Nigel Scrutton and colleagues extended this finding across aromatic amine dehydrogenase, morphinone reductase, and related enzymes, establishing tunneling as a general feature of this enzyme class rather than a localized curiosity.12
Crucially, this tunneling is conformationally gated. The protein matrix undergoes highly coordinated, large-scale dynamic fluctuations, on timescales from picoseconds to microseconds, that compress the donor-acceptor distance to within tunneling range, at which point the quantum-mechanical transfer occurs.13 The enzyme is not a static scaffold. It is a dynamically coupled oscillating system whose conformational motions are tuned to the exact quantum-mechanical requirements of the reaction it catalyzes. Remove these precise dynamics, by studying the enzyme at cryogenic temperatures or by introducing mutations that rigidify the protein, and tunneling efficiency drops sharply.14
This finding has a direct implication for the framework being developed here. The enzyme is not merely a static geometric template; it is a resonant acoustic structure. Its function depends on low-frequency, collective vibrational modes that thread energy from the protein’s outer surface directly into the active site. This is effectively a biological standing wave: a spatially distributed, temporally stable pattern of oscillation where every amino acid residue, cofactor, and substrate moves in a precise phase relationship to the entire macrostructure.
To see just how profoundly this standing wave alters our understanding of life, we have to look at how plants and bacteria harvest light. In classical physics, we used to think molecules in a cell moved like billiard balls, clumsily bumping into each other until they randomly found where they needed to go. But in 2007, a landmark study by researchers around Graham Fleming fundamentally challenged this view. Looking at a bacterial light-harvesting complex (the Fenna-Matthews-Olson complex), they discovered that when a photon of light hits, the energy doesn't hop around blindly. Instead, it behaves like a synchronized quantum wave, testing multiple molecular pathways at the exact same time to find the absolute fastest route to the reaction center.
There was just one catch: this hyper-efficient quantum behavior was initially observed at extreme, deep-freeze laboratory temperatures (−196∘C).15 Critics immediately argued that these delicate quantum tricks would instantly collapse into chaos in the warm, wet, and noisy environment of a living organism.
Yet just three years later, in 2010, Elisabetta Collini and her team stunned the physics community. They demonstrated that a species of marine cryptophyte algae achieves this exact same quantum synchrony at standard room temperature, proving that living systems are evolutionarily "wired" to sustain and harness quantum mechanics in everyday physiological conditions.16
The interpretation of this room-temperature phenomenon has been fiercely debated: are these purely electronic coherences, vibrational coherences, or vibronic coherences in which electronic quantum transitions are coupled to the rhythmic vibrations of the surrounding atomic nuclei? The emerging consensus favors the vibronic interpretation, and this matters. Vibronic coherences are far more robust against thermal noise than purely electronic ones, precisely because the mechanical motion of the protein matrix acts as a stabilizing cradle rather than a disruptive force.17
Though first mapped in photosynthesis, this principle applies broadly to dense, lipid-bound bioenergetic architectures. The cell is not a hot, incoherent molecular bath that suppresses quantum behavior. It is an environment that, at certain scales and within certain organized structures, actively sustains and exploits quantum coherence. The question is not whether coherence exists in biology; it does. The question is how far it extends, and what structural features support it.
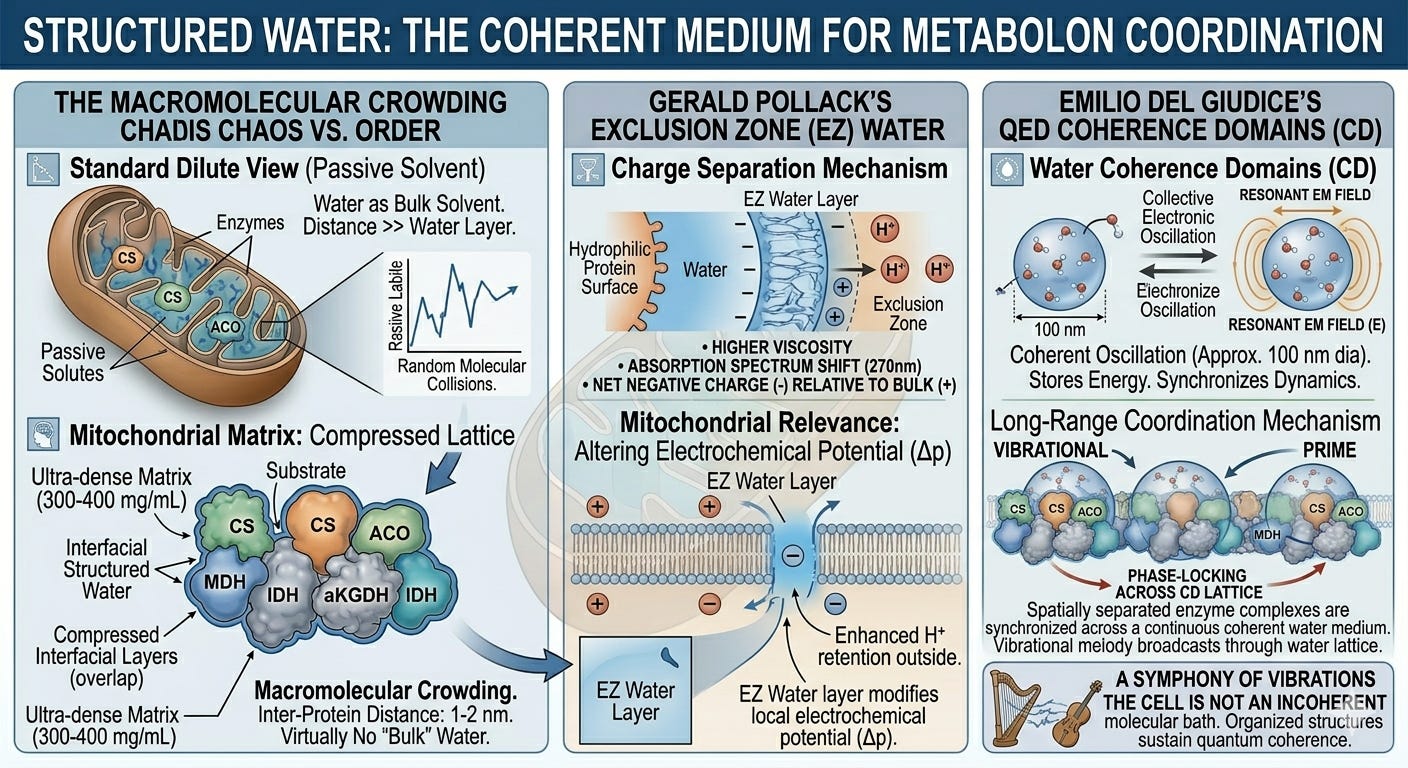
Structured Water: The Coherent Medium
Quantum coherence within individual enzyme complexes is striking. But the framework being developed here requires something more: a medium in which coherent dynamics can be coordinated across the assembled metabolon, and potentially across larger regions of the mitochondrial matrix. This is where the physical chemistry of cellular water becomes decisive, and where mainstream quantum biology has, until recently, lacked the analytical tools to fully map.
The standard biochemical treatment of water is as a passive solvent: a fluid background that merely dissolves cellular components. While that model works in a sparse, watery beaker, it completely fails inside the powerhouse of the cell. The mitochondrial matrix is the most crowded space in biology, packed with a protein density reaching up to 560 milligrams per milliliter.18 In this ultra-dense jungle, the average distance between proteins is a mere 1 to 2 nanometers. Because every single protein surface is wrapped in a jacket of structured, "exclusion-zone" water that extends outward, these water layers don't just touch, but are physically compressed together. Thermodynamically speaking, there is virtually no free "bulk water" inside a healthy mitochondrion. The matrix water is, to a first approximation, an unbroken, structured crystalline phase.19
Gerald Pollack’s work on exclusion zone (EZ) water established that water adjacent to hydrophilic surfaces forms an extended, liquid-crystalline phase with distinct physical properties: higher viscosity, a shifted absorption spectrum peaking around 270 nm, and a net negative charge relative to bulk water.20 This liquid-crystalline phase splits charge, pushing hydronium ions outward while retaining a localized negative charge at the surface.21 Inside a mitochondrion, which functions fundamentally as a proton-pumping battery, this intrinsic charge separation directly alters the local electrochemical potential adjacent to the inner membrane. The EZ water layer is not an idle spectator; it is a structural component of the bioenergetic circuit.
Emilio Del Giudice and colleagues developed a quantum electrodynamic (QED) account of water coherence that goes further still.22 On their analysis, water molecules in coherence domains, regions of roughly 100 nanometers in diameter under physiological conditions, oscillate collectively between two electronic states, coupled to a resonant electromagnetic field mode trapped within the domain.23 These coherent oscillations store energy, reduce the activation barriers for chemical reactions, and, most importantly, synchronize the dynamics of molecules embedded in or adjacent to the domain.
If Del Giudice’s coherence domains are sustained within this compressed matrix lattice, they provide the exact long-range coordination mechanism the metabolon requires. Rather than relying on blind molecular collisions, spatially separated enzyme complexes can be phase-locked across a continuous coherent medium. The vibrational activity of one enzyme broadcasts through the water lattice to prime the active site of the next.
This remains at the frontier of biophysics, and the QED account of water coherence is not yet mainstream. The structural evidence for highly organized interfacial water in the mitochondrial matrix is, however, well established, and the functional evidence that this organization matters for metabolic efficiency is growing.24 The QED interpretation of what that structure does remains contested; that the structure exists and is biologically consequential does not.
Counter-Space Geometry and the Standing Waveform
The evidence surveyed thus far: metabolon geometry, conformationally gated tunneling, vibronic coherence, and compressed structured water, converges on a picture deeply foreign to the linear, nodes-and-edges model of cell biology. It is a reality that the counter-space framework developed earlier in this series makes legible. Counter-space, as the geometry of orientation, relationship, and phase rather than mere position and distance, is the natural mathematical home for phenomena that are inherently non-local, oscillatory, and defined by their wholeness.
Each of the cellular mechanisms explored here shares these properties. Quantum tunneling is non-local in the precise sense that the proton or electron does not traverse the physical space between donor and acceptor; it occupies both simultaneously until the transfer is complete. Vibronic coherence is an emergent property of the entire system, not of any individual component; it cannot be isolated to a single molecule or bond. Exclusion zone water and Del Giudice coherence domains are spatially extended, phase-coherent entities whose defining characteristic is their collective oscillatory behavior. These are not point-space phenomena that happen to display interesting properties. They are counter-space, phase-field phenomena that manifest in point-space as the local condensations we call molecules, bonds, and reaction intermediates.
The Krebs cycle metabolon, on this reading, is a standing waveform. Each intermediate is not a distinct molecule waiting to be grabbed by the next enzyme; it is a node in a spatially distributed, temporally stable pattern of coupled oscillations. The cycle turns not because substrates diffuse sequentially across physical distance, but because the underlying waveform propagates through its counter-space phase space, each metabolic node reaching its characteristic vibrational state in rhythmic sequence, as in any standing wave. The molecular species that appear in the textbook diagrams are real, but they are the local, point-space condensations of the waveform’s nodes. They are the footprints; the waveform is the walker.
This reframing fundamentally redefines how the cycle can be disrupted and how it must be restored. A standing wave is disrupted not primarily by removing one of its nodes, since the wave will attempt to compensate by redistributing its amplitude, but by degrading the boundary conditions that sustain it, or by introducing incoherent noise that desynchronizes the phase relationships between nodes.
Mitochondrial dysfunction, on this view, is fundamentally a loss of coherence: a physical degradation of the liquid-crystalline water matrix and the precise phase relationships that constitute the bioenergetic waveform, rather than a simple deficiency of a single intermediate or cofactor. The clinical correlates are familiar to anyone practicing complex medicine: the diffuse, multi-system character of mitochondrial illness; the failure of single-nutrient interventions to restore long-term vitality; and the hyper-sensitivity of mitochondrial efficiency to factors such as membrane geometry, cardiolipin composition, and the cellular redox environment.25
Restoring the waveform requires restoring its coherent boundary conditions. In practical terms this means addressing the structural integrity of the mitochondrial inner membrane and its cristae architecture through targeted lipid replacement to protect and rebuild cardiolipin. It means restoring the interfacial EZ water environment of the matrix through thermodynamic inputs: photobiomodulation, infrared light, and grounding. It means clearing the oxidative, toxic, and inflammatory noise that desynchronizes protein dynamics. And at the level of the counter-space field, it means supporting the morphogenetic templates from which the molecular standing wave condenses in the first place.
Cofactor supplementation is not wrong. CoQ10 genuinely matters for electron transfer; NAD⁺ availability genuinely limits dehydrogenase function. But these interventions operate at the level of the point-space condensation: they adjust the amplitude at an isolated node. They do not address the waveform itself. A therapeutic approach adequate to the standing waveform model will use cofactor support as one element within a broader strategy aimed at coherence, geometry, and boundary conditions: the primary determinants of whether the waveform can sustain itself.
Images created using Google Gemini (Gemini 3.5 Flash).
References
For an overview of the shift from reconstituted to in-cell models of metabolism, see: Sweetlove LJ, Fernie AR. The role of dynamic enzyme assemblies and substrate channelling in metabolic regulation. Nat Commun. 2018;9:2136.
Pollack GH. The fourth phase of water: beyond solid, liquid, and vapor. Seattle (WA): Ebner & Sons; 2013.
Krebs HA, Johnson WA. Metabolism of ketonic acids in animal tissues. Biochem J. 1937 Apr;31(4):645–660.
Nicolson GL. Mitochondrial dysfunction and chronic disease: treatment with natural supplements. Integr Med (Encinitas). 2014 Aug;13(4):35–43.
Srere PA. The metabolon. Trends Biochem Sci. 1985 Mar;10(3):109–110.
Vélot C, Mixon MB, Teige M, Srere PA. Model of a quinary structure between Krebs TCA cycle enzymes: a model for the metabolon. Biochemistry. 1997 Nov 25;36(46):14271–14276.
Wheeldon I, Minteer SD, Banta S, Calabrese JC, Sarkar CA, Wu F. Substrate channeling as an approach to cascade biocatalysis. Nat Chem. 2016 Apr;8(4):299–309.
Bulutoglu B, Garcia KE, Wu F, Minteer SD, Banta S. Direct evidence for metabolon formation and substrate channeling in recombinant TCA cycle enzymes. ACS Chem Biol. 2016 Oct 21;11(10):2847–2853.
Schägger H, Pfeiffer K. Supercomplexes in the respiratory chains of yeast and mammalian mitochondria. EMBO J. 2000 Apr 17;19(8):1777–1783.
Ikon N, Ryan RO. Cardiolipin and mitochondrial cristae organization. Biochim Biophys Acta Biomembr. 2017 Jun;1859(6):1156–1163.
Klinman JP, Kohen A. Hydrogen tunneling links protein dynamics to enzyme catalysis. Annu Rev Biochem. 2013;82:471–496.
Scrutton NS, Hay S, Sutcliffe MJ. Quantum tunnelling in enzyme-catalysed hydrogen transfer: the 'ins and outs' of active-site dynamics. Biochem Soc Trans. 2002 Apr;37(Pt 2):328–335.
Hammes-Schiffer S, Stuchebrukhov AA. Theory of coupled electron and proton transfer reactions. Chem Rev. 2010 Dec 8;110(12):6939–6960.
New insight into quantum mechanical hydrogen tunneling in enzymes. Biochemistry. 2026 Feb 3;65(3):[Epub ahead of print].
Engel GS, Calhoun TR, Read EL, Ahn TK, Mancal T, Cheng YC, Blankenship RE, Fleming GR. Evidence for wavelike energy transfer through quantum coherence in photosynthetic systems. Nature. 2007 Apr 12;446(7137):782–786.
Collini E, Wong CY, Wilk KE, Curmi PM, Brumer P, Scholes GD. Coherently wired light-harvesting in photosynthetic marine algae at ambient temperature. Nature. 2010 Feb 4;463(7281):644–647.
Romero E, Augulis R, Novoderezhkin VI, Cogdell RJ, Gorrenz-Fraga AV, van Grondelle R. Quantum coherence in photosynthesis for efficient solar-energy conversion. Nat Phys. 2014 Sep;10(9):676–682.
Ellis RJ. Macromolecular crowding: obvious but underappreciated. Trends Biochem Sci. 2001 Oct 1;26(10):597–604.
Ellis RJ. Macromolecular crowding: obvious but underappreciated. Trends Biochem Sci. 2001 Oct 1;26(10):597–604.
Pollack GH. The fourth phase of water: beyond solid, liquid, and vapor. Seattle (WA): Ebner & Sons Publishers; 2013.
Zheng JM, Pollack GH. Long-range forces extending from polymer-gel surfaces. Phys Rev E Stat Nonlin Soft Matter Phys. 2003 Sep;68(3 Pt 1):031408.
Del Giudice E, Preparata G, Vitiello G. Water as a free electric dipole laser. Phys Rev Lett. 1988 Aug 29;61(9):1085–1088.
Del Giudice E, Vitiello G. Role of the electromagnetic field in the formation of domains in the process of symmetry-breaking phase transitions. Phys Rev A. 2006 Aug 17;74(2):022105.
Chaplin M. Do we underestimate the importance of water in cell biology? Nat Rev Mol Cell Biol. 2006 Nov;7(11):861–866.
Wallace DC. A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: a dawn for evolutionary medicine. Annu Rev Genet. 2005;39:359–407.